Die "Währungseinheit" für Energie innerhalb unseres Körpers ist das Molekül Adenosintriphosphat, kurz ATP. Es spielt eine zentrale Rolle in unserem Energiehaushalt und ist die universelle biochemische Batterie, in der die Energie aus den Nährstoffen gespeichert wird. Halbleer heißt diese Batterie ADP (Adenosindiphosphat).
Die gewonnene Energie (ΔG) wird genutzt, um endergone Vorgänge wie Biosynthesen, Bewegungs- und Transportprozesse anzutreiben. Lebende Systeme sind auf die kontinuierliche Zufuhr von freier Energie angewiesen. Sie dient drei Hauptzielen:
- der Muskelkontraktion und anderen zellulären Bewegungen (z. B. Flimmerschlag),
- dem aktiven Transport von Molekülen und Ionen,
- der Synthese von Biomolekülen aus einfacheren Stoffen.
Diese Prozesse halten lebende Systeme weit entfernt vom Gleichgewichtszustand. Bewegt sich eine Reaktion zum Gleichgewicht hin, dann wird Energie als Wärme frei und geht so dem System verloren. Die zentrale Bedeutung des ATP für den Energieaustausch in lebenden Systemen haben 1941 Fritz Lipmann und Herman Kalckar erkannt. Es ist in nahezu jeder Energieübertragung beteiligt.
ATP hat das Bestreben, durch Abspaltung der endständigen Phosphatgruppe eine „Entspannung“ zu erreichen. Bei diesem Prozess wird Energie frei. Diese Energiequelle nutzt die Zelle, indem sie mit Hilfe von Enzymen Phosphatgruppen des ATP auf andere Moleküle überträgt. Diese Moleküle können nun eine Arbeit verrichten, wobei die Phosphatgruppe wieder abgespalten wird. Die Delta G-Werte hängen von der Ionenkonzentration des Plasmas ab, z. B. den Konzentrationen von Mg2+ und Ca2+. Unter typischen zellulären Bedingungen beträgt der Wert ΔG = − 50 kJ/mol.
Soll Energie von einer exergonischen (energiefreisetzenden) auf eine endergonische (energieverbrauchende) Reaktion übertragen werden, so tritt ATP als Überträger auf. Die Energie, die bei der ATP-Hydrolyse freigesetzt wird, dient zum Antrieb für Reaktionen, die die Zufuhr von Energie erfordern, z. B. Muskelkontraktionen. Damit die Zelle weiterexistieren kann, muss sie ihren ATP-Vorrat immer wieder aus ADP und anorganischem Phosphat auffüllen. Eine aktive Muskelzelle regeneriert beispielsweise in jeder Sekunde ungefähr zehn Millionen ATP-Moleküle.
So entsteht umgekehrt ATP aus ADP und Pi, wenn Nährstoffmoleküle in chemoheterotrophen Organismen oxidiert werden oder wenn fotoautotrophe Organismen Licht einfangen:
ATP ⇄ ADP
Fotosynthese / Oxidation von Nährstoffmolekülen
Damit ist dieser ATP/ADP-Zyklus der fundamentale Mechanismus für den Energieaustausch in lebenden Systemen.
Energieumwandlung in der Zelle: Von Nahrungsbausteinen zu ATP
Wie wird die Energie, die im ATP gespeichert ist, aus den Nahrungsbausteinen gewonnen? Am Beispiel der Umwandlung der Glucose - der sogenannten Glykolyse - wollen wir das erklären: Im ersten Schritt wird das Glucose-Monosaccharid, ein Molekül mit sechs Kohlenstoffatomen (C-6-Körper), mithilfe von Enzymen in zwei C-3-Körper, das Pyruvat, gespalten. Die Glykolyse (= Zuckerzerlegung) läuft im Cytoplasma ab. Bei diesem Vorgang wird das Glucose-Molekül zunächst in zwei C3-Körper gespalten. Dies geschieht durch zweifache Phosphorylierung, also Abspaltung eines H und Anlagerung eines Phosphatrestes (P), so dass erst Glucose-6-Phosphat (C6H11O6P) entsteht (durch Strukturveränderung wird dieses in Fructose-6-Phosphat umgewandelt) und dann Fructose-1,6-bis-Phosphat (C6H10O6PP). Durch die Phosphorylierung ist Glucose nun im aktivierten Zustand. Dieser C6-Körper wird dann in ein Molekül Dihydroxyacetonphosphat (DHAP) und ein Molekül Glycerinaldehyd-3-Phosphat (GAP) (beide C3H5O3P) gespalten. Nur das Glycerinaldehyd-3-Phosphat kann weiterverwendet werden, deswegen wird das Dihydroxyacetonphosphat in dieses umgewandelt. Ein weiteres Phosphat wird angelagert und das Molekül wird oxidiert, wodurch 1,3-Bisphosphoglycerat (C3H4O4PP) entsteht. Die Elektronen werden auf den Wasserstoffüberträger NAD+ (Nicotinamidadenindinukleotid in der oxidierten Form) übertragen. Im weiteren Schritt wird ein Phosphatrest (P) auf ADP übertragen, so dass ATP und 3-Phosphoglycerinsäure (PGS, C3H5O4P) entstehen. Durch Abspaltung von Wasser entsteht nun Phosphoenolpyruvat (PEP, C3H3O3P).
Die Zellen bauen mit Hilfe ihrer Enzyme komplexe organische Moleküle ab (z. B. Glucose), die reich an Energie sind, und setzen sie in einfachere Produkte mit geringerem Energiegehalt um. Ein Teil der Energie kann Arbeit verrichten, der Rest geht als Wärme verloren.
Die Zellatmung ist ein Prozess, bei dem energiereiche in energiearme Stoffe abgebaut werden. Ziel der Zellatmung ist also die Energiegewinnung. Insgesamt werden bei der Zellatmung also energiereiche Stoffe abgebaut, damit diese freigewordene Energie dann von der Zelle genutzt werden kann. Bei dieser Umwandlung entsteht neben der Energie auch Wasser und Kohlenstoffdioxid. Diese Stoffe sind dann wieder die Grundbausteine für die Fotosynthese. Fotosynthese und Zellatmung sind ein zusammenhängender Kreislauf: Die Fotosynthese stellt die Ausgangsstoffe für die Zellatmung her, und die Endprodukte der Zellatmung sind die Ausgangsstoffe für die Fotosynthese.
Die Zellatmung lässt sich mit folgender Wortgleichung zusammenfassen:
Glucose + Sauerstoff → Kohlenstoffdioxid + Wasser + Energie
Als Zellatmung, biologische Oxidation oder innere Atmung werden jene Stoffwechselprozesse bezeichnet, die dem Energiegewinn der Zellen dienen. Insbesondere versteht man hierunter die biochemischen Vorgänge der Atmungskette in der inneren Membran der Mitochondrien, an deren Ende ATP synthetisiert wird.
Die Änderung der Freien Energie unter Standardbedingungen (Temperatur 25 °C, Druck 1,013 bar, Konzentration der an der Umsetzung beteiligten Stoffe 1 mol/L mit Ausnahme von Wasser und Gasen) beträgt bei der Oxidation von Glucose ΔG0' = - 2822 kJ je Mol Glucose. Bei biologischen Systemen wird für die H+-Ionen-Konzentration pH 7 statt pH 0 vereinbart, was die tatsächliche freie Energie beeinflusst.
Bei dieser chemischen Stoffumsetzung werden in einer Reihe von komplizierten Reaktionsschritten - darunter viele Redoxreaktionen - Wasserstoffatome von den Abbauprodukten der Glucose-Moleküle abgetrennt und mit Hilfe von Wasserstoffüberträgern (Nicotinamid-Adenin-Dinucleotid, NAD) zu den Mitochondrien transportiert. Dort reagieren die Wasserstoffatome innerhalb der Atmungskette mit Sauerstoff zu Wasser („biologische Knallgasreaktion“); die Glucose-Moleküle werden letztlich vollständig oxidiert. Am Ende des Abbauprozesses gewinnt die Zelle mit Hilfe der bei der biologischen Wasserstoff-Oxidation frei werdenden Energie die energiereiche Verbindung Adenosintriphosphat (ATP).
Der Abbau von einem Mol Glucose erbringt 35-38 Mol ATP. Pro ATP können 30 kJ Energie gerechnet werden: 38 * 30 kJ = 1140 kJ pro Mol Glucose. Der Wirkungsgrad der Zellatmung berechnet sich somit zu 1140 kJ / 2872 kJ = 0,39 oder 39 %.
Die Regulation der Zellatmung sorgt dafür, dass Energie nicht verschwendet, sondern z. B. in Form von chemischen Speicherstoffen wie Glucose, Stärke oder Fett gespeichert werden. Ist der Gehalt an ATP in den Zellen hoch, wird der Abbau verringert und stattdessen die Glukoneogenese oder die Glykogensynthese aktiviert. Regulationspunkte sind in den Prozessen der Zellatmung in Glykolyse, Citratzyklus und oxidativer Carboxylierung vorhanden. Oftmals funktioniert die Regulation über eine allosterische Aktivierung bzw. Inaktivierung der Enzyme.
Mitochondrien: Die Kraftwerke der Zelle
Mitochondrien sind die Zellorganellen, in denen bei Pflanzen und Tieren die Energiegewinnung stattfindet. Sie sind die Kraftwerke der Zelle. Hier wird die Glucose abgebaut, wodurch Energie entsteht. Die Mitochondrien sind Organellen mit doppelter Membran, die an der Produktion von ATP durch oxidative Phosphorylierung beteiligt sind. Sie besitzen ihre eigene DNA, was darauf hindeutet, dass sie früher eigenständige Organismen waren.
Der größte Teil zellulären ATPs entsteht durch oxidative Phosphorylierung in den Mitochondrien oder durch Fotophosphorylierung in den Chloroplasten. Weit weniger ATP wird über die Anlagerung/Übertragung anorganischer Phosphatreste von organischen Molekülen auf ADP erzeugt. Die oxidative Phosphorylierung ist allerdings von der Anwesenheit von Sauerstoff abhängig (aerobe Bedingungen).
Die Funktion der Mitochondrien besteht darin, als Kraftwerke der Zelle zu agieren. Sie sind der Hauptort der ATP-Produktion und optimieren Energieprozesse durch die Kombination mehrerer chemischer Reaktionen. Sie führen die oxidative Phosphorylierung durch, die in mehreren Schritten erfolgt: Elektronentransportkette zur Erzeugung eines Protonengradienten und Nutzung des Protonengradienten durch ATP-Synthase zur ATP-Produktion.
Die chemische Reaktion der ATP-Synthese in Mitochondrien lautet: \[ \text{ADP} + \text{P}_\text{i} + H^+ \rightarrow \text{ATP} + H_2O \].
Mitochondrien sind für die Zellatmung verantwortlich und liefern bis zu 95% der Energie, die eine Zelle benötigt. Ihre Funktion geht über die ATP-Produktion hinaus: Mitochondrien spielen eine Rolle bei der Regulation des Kalziumgleichgewichts, induzieren programmierte Zelltode (Apoptose) und sind an der Regulation der zellulären Signalgebung beteiligt.
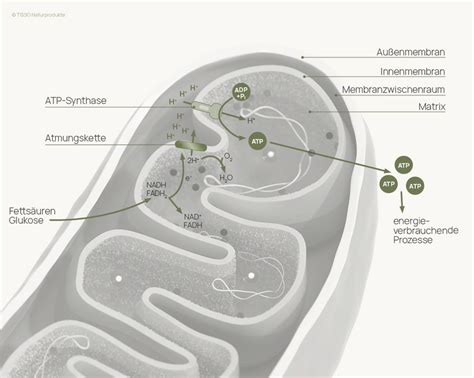
Der Citratzyklus und die Atmungskette
Beim Citratzyklus (auch Krebszyklus genannt) aus dem in der Glykolyse entstandenen Pyruvat entsteht CO2 und Essigsäure. Die Essigsäure durchläuft nun den Citratzyklus: Aus der Essigsäure und einem Oxalat aus vier Kohlenstoffatomen entsteht ein Citrat - das ist ein Molekül mit sechs Kohlenstoffatomen. Aus dem Citrat entstehen zwei CO2-Moleküle und ein Oxalat, das wiederum aus vier Kohlenstoffatomen besteht. Nun beginnt der Kreislauf wieder von vorn: Aus Oxalat und Essigsäure wird ein Citrat, aus dem Citrat werden wieder zwei CO2-Moleküle und ein Oxalat und so weiter. Das entstehende CO2 atmen wir aus.
Die Atmungskette befindet sich in der inneren Mitochondrienmembran. Sie spielt eine zentrale Rolle in der zellulären Energiegewinnung durch die Übertragung von Elektronen entlang von Proteinkomplexen. In den Mitochondrien wird der von den NADH-Molekülen bereitgestellte Wasserstoff über eine Kette von Enzymen weitertransportiert. Von Enzym zu Enzym wird frei werdende Energie in ATP-Molekülen gespeichert: Der Atmungskette wird stufenweise Energie entzogen. Erst am Ende der Kette läuft eine Reaktion von Wasserstoff mit Sauerstoff ab, als Abfallprodukt entsteht dabei Wasser.
Ein NADH kann 2 Elektronen (e−) abgeben, wodurch der am NAD gebundene Wasserstoff als Proton (H+) frei wird und das verbleibende NAD-Molekül positiv geladen wird: NAD+. Weil die so abgegebenen 2 Elektronen auf einem recht hohen Energieniveau liegen (sehr niedriges Redoxpotential des Redoxpaares NADH/NAD+), können mit ihrer Hilfe 10 Protonen aus der Matrix in den Intermembranraum transportiert werden. Dies geschieht folgendermaßen: Die 2 Elektronen des NADH reduzieren den ersten Komplex (Komplex I) von mehreren Enzym-Komplexen der Atmungskette, die sich zwischen Matrix und Intermembranraum des Mitochondriums befinden. Jedes Elektron wird nun über Redoxreaktionen von einem Enzym-Komplex zum nächsten weitergegeben. Aufgrund der Weitergabe von Elektronen von Komplex zu Komplex wird dieser Vorgang auch als Elektronentransportkette bezeichnet.
Durch den Komplex I, den Komplex III und den Komplex IV werden H+-Ionen (Protonen) aus der Matrix in den Intermembranraum transloziert. Im Intermembranraum entsteht auf diese Weise eine hohe Wasserstoffionen-Konzentration, wodurch ein pH-Wert unter 7 entsteht, und es entsteht ein osmotisches Potential. Die Wasserstoffionen fließen schließlich durch die membrangebundene ATP-Synthase vom Intermembranraum in den Matrixraum zurück. Die Durchflussenergie wird dazu genutzt, einen Phosphatrest an ADP zu binden. Durch die Oxidation von einem NADH entstehen so 3 ATP. Ausnahme sind die zwei NADH aus der Glykolyse. Diese befinden sich noch im Cytoplasma und müssen erst in die Mitochondrien transportiert werden, daher gewinnt man aus diesen nur je 2 ATP. Da 8 + 2 NADH oxidiert werden, entstehen insgesamt 8 × 3 + 2 × 2 = 28 ATP.
Mit dem FADH2 funktioniert der Vorgang im Prinzip genauso, nur gibt FADH2 auf einem höheren Redoxpotential, also niedrigeren Energieniveau, Elektronen ab und deshalb erst auf einer energetisch niedriger stehenden Stufe in die Atmungskette ein. Deshalb können mit Hilfe der Elektronen des FADH2 nur 4 Protonen aus der Matrix in den Intermembranraum gepumpt werden. Mit einem FADH2 werden darum nur 2 ATP gebildet. Die Protonen und die Elektronen des NADH und des FADH2 (jeweils insgesamt 24) werden zusammen mit 6 O2, die durch die Membran in die Mitochondrienmatrix transportiert werden, zu 12 H2O oxidiert. Die Elektronen- bzw. Wasserstoffüberträger NAD+ und FAD können durch Aufnahme von je 2 e− und 2 H+ wieder zu NADH bzw. FADH2 reduziert werden.
Der Zellzyklus: Wachstum, Teilung und Differenzierung
Wenn ihr euch in der Schule gerade mit Zellen und deren Aufbau beschäftigt, ist bestimmt auch die Zellatmung Teil des Unterrichts. Doch wie atmet eine Zelle? Welche Vorgänge laufen wie genau dabei ab und welche Produkte kommen dabei heraus? Zellatmung einfach erklärt: Bei der Fotosynthese wird aus Licht und Kohlenstoffdioxid Sauerstoff und Glucose (Traubenzucker) hergestellt. Der Zucker, der hier entsteht, wird bei der Zellatmung verbraucht, um Energie für die Pflanze zu gewinnen. Ziel der Zellatmung ist also die Energiegewinnung. Sie findet in den sogenannten Mitochondrien der Zelle statt.
Die Zelle wird im Allgemeinen aus thermodynamischer Sicht als offenes System bezeichnet. Das offene System betreibt mit seiner Umgebung Stoffaustausch und Energieaustausch. In dem System Zelle werden ständig Produkte entzogen und notwendige Edukte zugeführt. Durch den ständigen Zufluss bzw. Abfluss von Molekülen und damit Energie kann sich kein chemisches Gleichgewicht einstellen. Erst mit dem Tode kommt dieser Prozess zum Stillstand.
Merke: Zelle: offen für Energie- und Stoffaustausch.
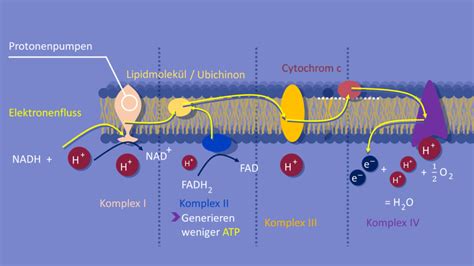
Phasen des Zellzyklus
Der Zellzyklus beschreibt die Abfolge von Ereignissen, die eine Zelle durchläuft, um sich zu teilen. Er besteht aus verschiedenen Phasen:
Interphase
Die längste Phase des Zellzyklus ist die Interphase, die sich in drei Unterphasen gliedert:
- G1-Phase (Gap 1): Nach der Zellteilung beginnt die G1-Phase. Das "G" steht für "gap", auf Deutsch so viel wie "Lücke". In dieser Phase wächst die Zelle und synthetisiert Proteine und Organellen. Die meisten Zellen eines vielzelligen Eukaryoten teilen sich nicht ewig, sondern spezialisieren sich zu Gewebezellen mit definierten Aufgaben. Diese Spezialisierung von Zellen auf bestimmte Aufgaben wird als Zelldifferenzierung bezeichnet. Differenzierte Zellen verlieren normalerweise ihre Fähigkeit zur Zellteilung und verharren in einer aktiven Phase, die als G0-Phase bezeichnet wird. Unter bestimmten Umständen, z. B. nach Verletzungen, besteht die Notwendigkeit zur Regeneration von Geweben, was dazu führen kann, dass differenzierte Zellen die Fähigkeit zu erneuten Mitosen erhalten (Reembryonalisierung).
- S-Phase (Synthese): In der S-Phase findet die Replikation der DNA statt. Aus den 1-Chromatid-Chromosomen werden wieder 2-Chromatid-Chromosomen, die jeweils zwei identische DNA-Fäden enthalten. Neben der DNA müssen natürlich auch viele Proteine hergestellt werden, vor allem die Histone, die für die "Verpackung" der DNA-Doppelhelix wichtig sind.
- G2-Phase (Gap 2): An die S-Phase schließt sich wieder eine "gap"-Phase an, die G2-Phase. Hier wird die nächste Mitose vorbereitet, falls überhaupt noch eine stattfinden soll. Dazu werden RNA- und Protein-Moleküle synthetisiert, die für die kommende Mitose wichtig sind.
M-Phase (Mitose und Cytokinese)
Die M-Phase umfasst die eigentliche Zellteilung:
- Mitose: Bei der Mitose wird das genetische Material der Mutterzelle gleichmäßig auf zwei Tochterzellen aufgeteilt. Dazu werden die 2-Chromatid-Chromosomen in ihre jeweils zwei Chromatiden aufgespalten und dann wird jeweils das eine Chromatid in die eine Tochterzelle, das andere, genetisch identische Chromatid in die andere Tochterzelle verfrachtet. Zu Beginn der Mitose (M-Phase) wird das mitochondriale Protein Tom6 phosphoryliert, was zu einem verstärkten Import von Proteinen führt, die die Kraftwerksleistung der Mitochondrien verbessern. Dies stellt sicher, dass für ein aufwändiges Ereignis wie die Zellteilung genügend Energie vorhanden ist.
- Cytokinese: Dies ist die Teilung des Zytoplasmas. In Tierzellen wird während der späten Anaphase und in der Telophase der Mitose ein Ring aus Aktin- und Myosinfasern gebildet, zwischen den beiden künftigen Tochterzellen. Ähnlich wie bei den Vorgängen in Muskelzellen zieht sich dieser Actin-Myosin-Ring unter ATP-Verbrauch zusammen, und die Zelle verengt sich in der Mitte. In Pflanzenzellen wird während der Telophase in der Äquatorialebene ein Phragmoplast gebildet.
Kontrollpunkte des Zellzyklus
Unter den Kontrollpunkten des Zellzyklus versteht man bestimmte Stadien oder Zeitpunkte des Zyklus, an denen die Zelle überprüft, ob die nächste Phase des Zyklus eingeleitet werden kann. Zahlreiche Schlüsselenzyme sind bei dieser Kontrolle des Zellzyklus und damit bei der Kontrolle des Zell- und Gewebewachstums beteiligt.
Am Ende der G1-Phase befindet sich ein wichtiger Kontrollpunkt. Wird dieser Kontrollpunkt ungehindert überschritten, dann folgt unweigerlich die nächste Mitose und die nächste Zellteilung. Wenn dieser Kontrollpunkt dagegen nicht überschritten wird, gelangt die Zelle in die G0-Phase. Zumindest bei Hefezellen ist sicher, dass der Kontrollpunkt 1 durch Außenfaktoren wie Nährstoffangebot und Pheromone sowie durch die Zellgröße beeinflusst werden kann.
Sollte es aus irgendeinem Grund bei der Replikation der DNA zu größeren Problemen gekommen sein, wird der Zellzyklus an dieser Stelle angehalten. Auch bei der Mitose können Fehler passieren. In diesem Fall wird der Zellzyklus ebenfalls angehalten.
Zwei Familien von Schlüsselproteinen spielen eine wichtige Rolle bei der Kontrolle des Zellzyklus: die Cycline und die cyclinabhängigen Proteinkinasen. Cycline sind "stadienspezifisch im Zellzyklus auftretende Proteine, die cyclinabhängige Proteinkinasen aktivieren können". Nachdem die Cycline ihre Aufgabe erfüllt haben, werden sie sofort abgebaut, damit nicht die gleiche Phase des Zellzyklus zweimal abläuft. Die cyclinabhängigen Proteinkinasen können auch von bestimmten Phosphatasen wie zum Beispiel Cdc25 phosphoryliert und damit aktiviert werden. Auch gibt es hemmende Enzyme (ICK = inhibitor of cyclin-dependet kinase), die diese cyclinabhängigen Proteinkinasen vorübergehend stilllegen.
Zelluläre Energiegewinnung und Zellteilung: Eine Verknüpfung
Mitochondrien liefern als Kraftwerke der Zelle die Hauptenergie für die Prozesse des Zellzyklus. Sie wandeln Nahrung in Energie um, sodass die Zelle sie nutzen kann. Die Freiburger Biochemikerin Dr. Angelika Harbauer und der Biochemieprofessor Chris Meisinger haben zusammen mit einem deutsch-französischen Team einen Signalweg entdeckt, der die beiden zentralen Aufgaben Zellteilung und Energieumwandlung miteinander verknüpft.
Bei der Zellteilung verdoppelt sich die Erbinformation und verteilt sich in einem komplexen Prozess, den man Mitose nennt, auf die entstehenden Tochterzellen. Bestimmte Eiweiße, so genannte Cyclin-abhängige Kinasen, steuern diese Prozesse. Wie ihr Name andeutet, werden die Eiweiße von Cyclinen reguliert: Diese sind andere Proteine, die während der verschiedenen Zellzyklusphasen periodisch auf- und wieder abgebaut werden.
Harbauer hat am Modell der Bäckerhefe herausgefunden, dass zu Beginn der Mitose eine Cyclin-abhängige Kinase ein mitochondriales Protein phosphoryliert und dadurch aktiviert. Dieses mitochondriale Protein ist ein Bestandteil der Proteineintrittspforte der Mitochondrien, die nahezu alle Proteine importiert, die in den Mitochondrien eine Aufgabe erfüllen. Durch diese zellzyklus-abhängige Veränderung der Proteineintrittspforte kommt es zu einem verstärkten Import von Proteinen, die die Kraftwerksleistung der Mitochondrien verbessern. Dadurch ist sichergestellt, dass für ein aufwändiges Ereignis wie die Zellteilung genügend Energie vorhanden ist. Kann diese Veränderung an der Proteineintrittspforte nicht mehr stattfinden, verlangsamt das die Zellteilung und letztlich das Wachstum.
„Die in der Hefe entdeckten Modifikationen finden wir auch beim Menschen. Es erscheint daher sehr wahrscheinlich, dass dieser neue Signalweg auch dort die Zellkraftwerksleistung und die Zellteilung miteinander verbindet“, sagt Meisinger. Bei vielen Tumorerkrankungen sei die Leistung der Zellkraftwerke stark vermindert. Der entdeckte Signalweg könnte ein Schlüssel sein, den Zusammenhang zwischen fehlregulierter Zellteilung und den Mitochondrien aufzuklären.
Zellatmung - Übersicht
Die Dauer eines Zellzyklus ist von Zellart zu Zellart unterschiedlich, meistens handelt es sich jedoch nur um sechs bis 24 Stunden. Bei einem 16-Stunden Zellzyklus dauert die Mitose ca. 1 Stunde, die G1-Phase ca. 5 Stunden, die S-Phase ca. 7 Stunden und die G2-Phase ca. 3 Stunden.