Die Photosynthese, abgeleitet vom Altgriechischen φῶς (phō̂s) für „Licht“ und σύνθεσις (sýnthesis) für „Zusammensetzung“, ist ein fundamentaler physiologischer Prozess, der es Pflanzen, Algen und bestimmten Bakterien ermöglicht, energiereiche Biomoleküle aus energieärmeren Stoffen mithilfe von Lichtenergie zu erzeugen. Die Effizienz, mit der die eingestrahlte Lichtenergie für den Aufbau von Assimilaten genutzt wird, wird als photosynthetische Effizienz bezeichnet. Man unterscheidet dabei zwischen der oxygenen und der anoxygenen Photosynthese. Bei der oxygenen Form wird molekularer Sauerstoff (O2) freigesetzt.
Die Photosynthese stellt den wichtigsten biochemischen Prozess dar, bei dem Lichtenergie, meist Sonnenlicht, in chemisch gebundene Energie umgewandelt wird (Phototrophie). Fast alle heterotrophen Lebewesen sind indirekt von diesem Prozess abhängig, da sie letztlich ihre Nahrung und den für die aerobe Atmung notwendigen Sauerstoff der Photosynthese verdanken.
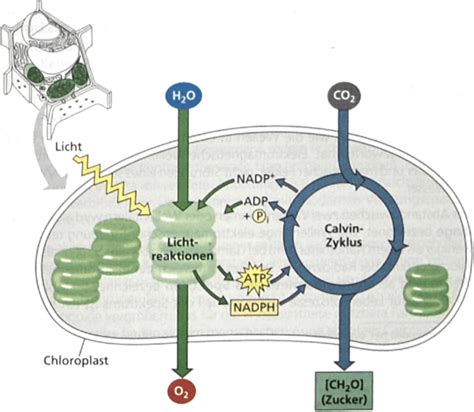
Die Licht- und Dunkelreaktionen der Photosynthese
Die ersten beiden Schritte der Photosynthese werden als Lichtreaktion bezeichnet und laufen bei Pflanzen in den Photosystemen I und Photosystem II ab. Die Synthese energiereicher organischer Stoffe geht überwiegend von der Kohlenstoffverbindung Kohlenstoffdioxid (CO2) aus. Für die Verwertung von CO2 muss dieses reduziert werden. Als Reduktionsmittel (Reduktans) dienen Elektronen aus oxidierbaren Stoffen wie Wasser (H2O), elementarem Wasserstoff (H2), Schwefelwasserstoff (H2S), zweiwertigen Eisenionen (Fe2+) oder einfachen organischen Stoffen wie Säuren und Alkoholen (z. B. Acetat bzw. Ethanol). Auch die Oxidation einfacher Kohlenhydrate kann Elektronen liefern.
Die allgemeine Formulierung H2A steht hierbei für das Reduktans, wobei H2O oder andere Verbindungen gemeint sind. Alle Algen und grünen Landpflanzen nutzen ausschließlich Wasser (H2O) als Reduktans H2A, wobei A für den im Wasser gebundenen Sauerstoff steht. Dieser wird bei der oxygenen Photosynthese als elementarer, molekularer Sauerstoff (O2) freigesetzt.
Photosynthetische Bakterien können eine größere Bandbreite an Reduktantien nutzen, vorwiegend Schwefelwasserstoff (H2S). In diesem Fall steht A für den im Schwefelwasserstoff gebundenen Schwefel, und es wird elementarer Schwefel (S) anstelle von Sauerstoff freigesetzt. Allen Formen der Photosynthese ist gemein, dass durch die Oxidation von Reduktantien Elektronen gewonnen werden. Der für die Synthese energiereicher organischer Verbindungen benötigte Kohlenstoff kann aus CO2 (Photoautotrophie) oder aus einfachen organischen Verbindungen gewonnen werden. Photoautotrophe Organismen, wie grüne Pflanzen und Algen, bilden das primäre Syntheseprodukt als phosphorylierte Triose, die als Ausgangsmaterial für den Aufbau von Bau- und Reservestoffen dient.
Historische Entwicklung des Verständnisses der Photosynthese
Seit der Antike herrschte die Vorstellung, dass Pflanzen ihre Nahrung aus der Erde beziehen. Erst Marcello Malpighi überprüfte 1671 experimentell, dass der Nahrungssaft in den Blättern durch Sonnenlicht verarbeitet wird. Jan Ingenhousz zeigte 1779, dass grünen Blättern bei Lichteinwirkung Sauerstoff gebildet wird. Die Humustheorie hielt sich jedoch bis Mitte des 19. Jahrhunderts, bis Justus von Liebigs Arbeiten zur Assimilation anorganischer Substanzen durch Pflanzen unabweisbar wurden.
Frederick Blackman und Gabrielle Matthaei postulierten 1905 die Unterscheidung zwischen einer photochemischen Lichtreaktion und einer enzymatischen Dunkelreaktion. Cornelis Bernardus van Niel schlug 1930 vor, dass Photosynthese ein Austausch von Wasserstoff zwischen einem Donator und Kohlendioxid als Akzeptor sei, wobei Wasser als Donator fungiert. Robert Hill lieferte 1937 einen Beleg dafür, indem er zeigte, dass isolierte Chloroplasten auch ohne CO2 Sauerstoff bilden, wenn Eisensalze als künstliche Elektronenakzeptoren vorhanden sind (Hill-Reaktion).
Lichtabsorption und Lichtsammelkomplexe
Die Energie des Lichts wird bei phototrophen Organismen durch Farbstoffe, sogenannte Chromophore, eingefangen. In grünen Pflanzen und Cyanobakterien sind dies Chlorophylle, in anderen Bakterien Bakteriochlorophylle. Diese absorbieren Licht unterschiedlicher Wellenlängenbereiche, wobei die charakteristische grüne Farbe durch die sogenannte Grünlücke entsteht.
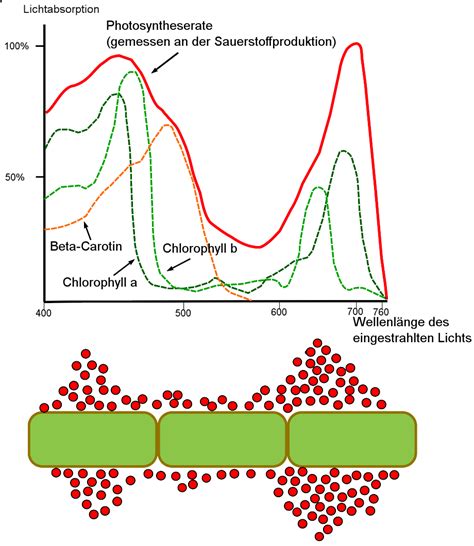
Wenn Licht auf ein Pigment trifft, geht das Chromophor in einen angeregten Zustand über. Die Energie für diese Anregung und damit das Absorptionsspektrum hängen vom Aufbau der konjugierten Doppelbindungen ab. Chlorophylle a und b absorbieren hauptsächlich blaues und rotes Licht, grünes Licht jedoch kaum. Das angeregte Chlorophyll kann ein Elektron auf einen Akzeptor übertragen, wobei ein positiv geladenes Chlorophyllradikal (Chl•+) zurückbleibt.
Die Anordnung von chlorophyllhaltigen Lichtsammelkomplexen zu Antennen um ein gemeinsames Reaktionszentrum vergrößert den Querschnitt und verbreitert das Absorptionsspektrum. Die eng benachbarten Chromophore in den Antennen leiten die Lichtenergie als Exziton weiter, bis sie das Reaktionszentrum erreicht. In Pflanzen bilden die Lichtsammelkomplexe eine Zentral- und eine äußere Antenne, eingebettet in die Thylakoidmembran. Carotine und Xanthophylle dienen als zusätzliche Chromophore und schützen die Antenne vor schädlichen Zuständen.
Photosysteme und der Elektronentransport
Bei allen grünen Pflanzen, Algen und Cyanobakterien sind die Photosysteme II und I funktionell hintereinander geschaltet. Sie nutzen Lichtenergie zur Speicherung von Adenosintriphosphat (ATP) und zur Gewinnung von Elektronen aus Wasser. Die Oxidation von Wasser führt zur Freisetzung von molekularem Sauerstoff (O2) und Protonen (H+) - der Photolyse des Wassers. Während dieses Vorgangs werden Protonen in das Lumen der Thylakoide transportiert.
Um Wasser oxidieren und NADP+ reduzieren zu können, sind zwei Photosysteme mit unterschiedlichen Redoxpotentialen erforderlich. Photosystem II stellt ein starkes Oxidationsmittel zur Wasserspaltung bereit, während an Photosystem I ein starkes Reduktionsmittel zur Reduktion von NADP+ erzeugt wird. Das bei der Lichtreaktion gewonnene Reduktionsmittel (NADPH) und die Energiequelle (ATP) werden im Calvin-Zyklus („Dunkelreaktion“) zur Synthese von Kohlenhydraten aus Kohlenstoffdioxid verwendet. Der freigesetzte Sauerstoff stammt aus Wasser, nicht aus CO2.
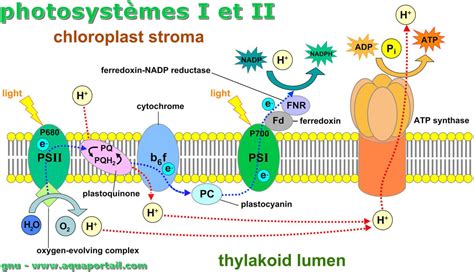
Die beiden Photosysteme sind durch eine Elektronentransportkette verbunden, die Plastochinon, den Cytochrom-b6f-Komplex und Plastocyanin umfasst. Das Zick-Zack-Diagramm der Redoxpotentiale verdeutlicht den Elektronentransfer.
Bei der Absorption eines roten Lichtquants wechselt das Chlorophyll-Molekül in den angeregten Zustand S1, bei blauem Licht in den energiereicheren Zustand S2. Nur S1 ist für die Photochemie nutzbar. Über strahlungslosen Energietransfer gelangen Exzitone zu den Reaktionszentren.
In den offenen Reaktionszentren von Photosystem II (P680) wird eine Ladungstrennung durchgeführt, bei der ein Elektron auf einen primären Akzeptor (Phäophytin) übertragen wird. Dieses Elektron durchläuft eine Kette von Akzeptoren, einschließlich Plastochinon (QB), das nach Aufnahme von zwei Elektronen und Protonen als Plastochinol (PQH2) in die Membranmatrix freigesetzt wird.
Das stark oxidierende P680•+-Radikal wird durch einen Tyrosinrest reduziert. PQH2 gelangt zum Cytochrom-b6f-Komplex, wo es durch das Rieske-Protein und Cytochrom-f weitergegeben wird. Das Elektron gelangt schließlich auf das reduzierte Plastocyanin (PC), ein kupferhaltiges Protein, das im Lumen des Thylakoiden diffundiert.
Das reduzierte Plastocyanin erreicht Photosystem I (PS I), dessen Chlorophyll-a-Paar (P700) durch Lichtabsorption in einen energetisch höheren Zustand versetzt und ein Elektron abgibt. Dieses Elektron wird auf einen ersten Akzeptor (A0) übertragen.
Regulation und Schutzmechanismen der Photosynthese
Die Photosynthese ist anfällig für Schäden durch reaktive Sauerstoffspezies (ROS), die insbesondere bei hoher Lichtintensität entstehen können. Diese ROS, wie Singulett-Sauerstoff (1O2), Superoxidradikale (O2*-) und Hydroxylradikale (OH*-), können Zellkomponenten schädigen.
Pflanzen verfügen über ausgeklügelte Schutzsysteme, um diese ROS zu neutralisieren. Dazu gehören antioxidative Moleküle wie Ascorbinsäure (Vitamin C) und Polyphenole sowie Enzyme wie Ascorbinsäure-Peroxidase.
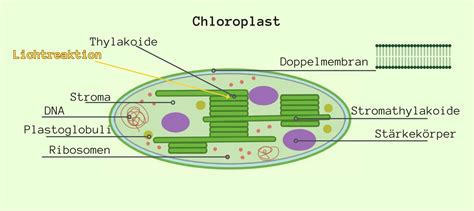
Im Ascorbat-Glutathion-Zyklus, der im Stroma der Chloroplasten stattfindet, wird Ascorbinsäure durch Ascorbinsäure-Peroxidase regeneriert, wobei ROS zu Wasser reduziert werden. Glutathion spielt ebenfalls eine wichtige Rolle bei der Entgiftung von ROS.
Ein weiterer wichtiger Schutzmechanismus ist das Non-photochemical quenching (NPQ), die energieabhängige Dissipation von überschüssiger Lichtenergie als Wärme. Dies wird durch Änderungen des Thylakoid-pH-Wertes und die Synthese von Zeaxanthin vermittelt.
Studien mit Mais-Chloroplasten zeigten, dass NPQ schnell reversibel ist und durch Ascorbinsäure beeinflusst wird. Hohe Konzentrationen von Ascorbinsäure verhinderten einen transienten Abfall von NPQ. Bedingungen, die die Ascorbat-abhängige Mehler-Peroxidase-Reaktion begünstigen, führten zu einer veränderten NPQ-Dynamik.
Schäden an der Photosyntheseapparatur, wie beeinträchtigter Elektronentransport oder Schäden an Enzymen wie Rubisco, können auftreten, wenn die Schutzsysteme überlastet sind. Die Fähigkeit, ROS abzuwehren und überschüssige Lichtenergie zu dissipieren, ist entscheidend für die Gesunderhaltung und Funktionsfähigkeit der photosynthetisch aktiven Zellen.